
Druck-Version
Evolution der Vierbeiner
Die evolutionäre Zwischenform Tiktaalik roseae
Zeuge eines berühmten Übergangs
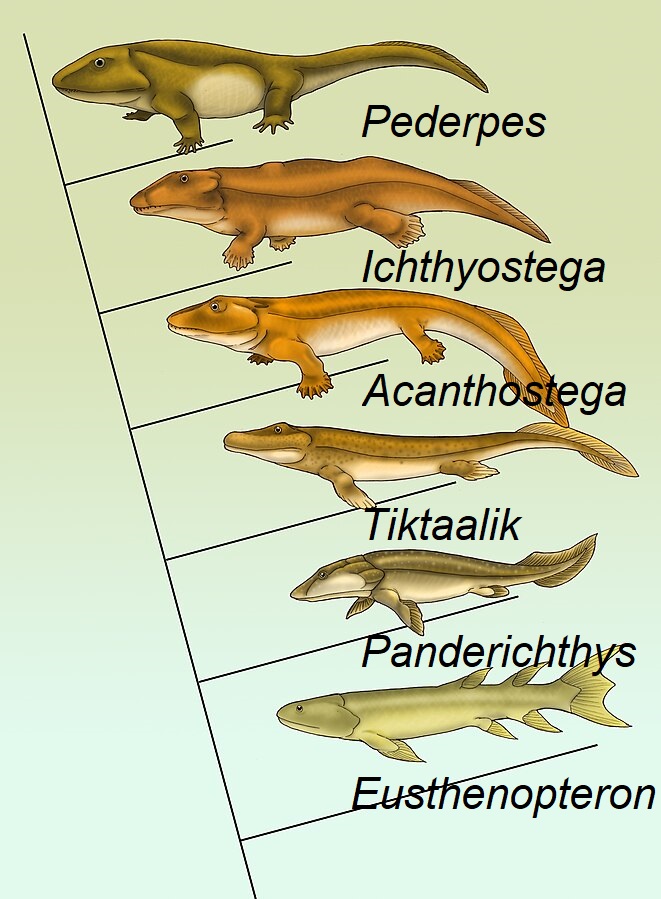
Die Entstehung der Landwirbeltiere (Tetrapoden) repräsentiert den wohl berühmtesten Übergang von der aquatischen Lebensweise zu einem Leben an Land. Diese Evolution lässt sich anhand fossil erhaltener Stammtetrapoden wie Eusthenopteron, Panderichthys und Acanthostega rekonstruieren; die Vierbeiner gingen also aus urtümlichen Fleischflossern (Sarcopterygiern) hervor. Dieser Entwicklungsabschnitt ist durch eine Reihe von Umwandlungen gekennzeichnet. Dazu zählt die Umstrukturierung von Schädel, Schultergürtel und der Region, aus der sich später das Mittelohr bildete. Zeitgleich entstanden aus Flossen robuste Extremitäten, die sich in Oberarm, Elle und Speiche des Unterarms, Handwurzel und Finger untergliedern.
Titelbild: Kladogramm ausgewählter Stammtetrapoden, der Vorfahren der vierbeinigen Landwirbeltiere. Bildquelle: Maija Karala, Fishapod evolution, Artbezeichnungen eingefügt, CC BY-SA 3.0.
{kind=link}
Lange wurde die Entstehung der Landwirbeltiere spärlich durch den Fossilienbefund veranschaulicht. So weisen die aquatisch lebenden Fleischflosser Eusthenopteron und Panderichthys noch wenige Tetrapoden-Neuerungen auf, während sich die amphibienähnlichen Fleischflosser Acanthostega und Ichthyostega anatomisch bereits deutlich von ihnen absetzen. Doch im Jahr 2006 berichtete die Wissenschaftszeitschrift Nature gleich in drei Artikeln über eine spektakuläre Neuentdeckung: den Raubfisch Tiktaalik roseae.
Die Bedeutung von Tiktaalik als evolutionäre Zwischenform
Tiktaalik (Abb. 1) verkörpert eine Zwischenform, die in die oben genannte morphologische Lücke fällt: Der Fisch mit dem krokodilähnlichen Schädel ähnelt einem Tetrapoden bereits mehr als die aquatischen Fleischflosser Panderichthys und Eusthenopteron, aber weniger als die amphibienähnlichen Gattungen Acanthostega und Ichthyostega. Anders gesagt, Tiktaalik zweigt im Kladogramm "zwischen" Panderichthys und Acanthostega von der Stammlinie der Vierbeiner ab (vgl. AHLBERG & CLACK 2006, S. 748 sowie Abb. 2).

Abb. 1 Computergestützte 3D-Rekonstruktion von Tiktaalik anhand mehrerer fossiler Exemplare. Das Tier ähnelte den Quastenflossern und Lungenfischen in der Beschuppung, im Bau von Flossen, Unterkiefer und Gaumen. Amphibienartig sind das verkürzte Schädeldach und die Ohrregion, der bewegliche Hals und die Vordergliedmaßen. Ausgestattet mit Ellenbogen- und Handgelenk erinnern die Brustflossen an Arme, sie endeten jedoch in Flossenstrahlen, nicht in Fingern. Bild: Daniel Mortell /Depositphotos.com.
Überreste mehrerer Exemplare wurden aus dem Schlick eines eingetrockneten Flussbetts auf der kanadischen Insel Ellesmere im Nunavut-Territorium, 1000 Kilometer vom Nordpol entfernt, geborgen. Zu ihren Entdeckern zählen unter anderem Edward DAESCHLER von der Academy of Natural Sciences (Philadelphia), Neil SHUBIN von der Universität Chicago (Illinois) und Farish JENKINS von der Harvard Universität in Cambridge.
Das Tier lebte im Ober-Devon vor 380 Millionen Jahren und verfügt über Flossen, die den Beginn der Entstehung der Gliedmaßen dokumentieren. Die Extremität zeigt Oberarmknochen, Elle, Speiche und Handwurzelknochen, weist aber noch keine eindeutigen Finger auf (DAESCHLER et al. 2006; SHUBIN et al. 2006). Des Weiteren zeigt das verkürzte Schädeldach, das Fehlen von Kiemendeckeln, die Schultergürtel und weitere Merkmale Anklänge an die Tetrapoden, während andere Merkmale wie die Flossenstrahlen ursprünglich blieben.
Tiktaalik passt nicht nur aufgrund seines Körperbaus in das zu erwartende Übergangsfeld zwischen aquatischen Fleischflossern und Tetrapoden. Auch stratigraphisch fällt Tiktaalik in die evolutionstheoretisch vorhergesagte Formation des untersten Ober-Devons (AHLBERG & CLACK 2006, S. 748). Tiktaalik ist damit 3 Mio. Jahre jünger als Panderichthys und 20 Mio. Jahre älter als die auf 365 Mio. Jahre datierten Überreste der Urtetrapoden Acanthostega und Ichthyostega. Das Tier wird von den Paläontologen als "echtes Bindeglied" eingestuft, da es dazu beiträgt, eine Lücke in unserem Verständnis darüber zu schließen, in welchen Etappen sich der Landgang der Wirbeltiere vollzog (DALTON 2006).
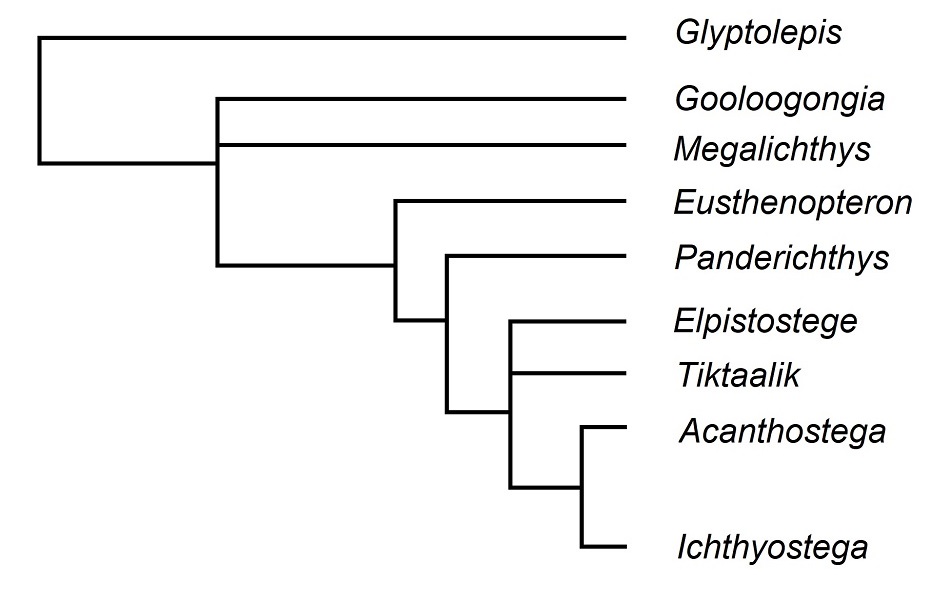
Abb. 2 Die phylogenetische Analyse von 114 Merkmalen und neun Sarcopterygier-Gruppen liefert folgendes Kladogramm als plausibelste Verwandtschaftshypothese ("consensus tree"). Tiktaalik verkörpert die nächstverwandte Gruppe (Schwestergruppe) von Acanthostega und Ichthyostega (Tetrapoden) und teilt diese Position mit Elpistostege. Panderichthys, Tiktaalik, Acanthostega usw. sind insofern evolutionsbiologisch relevante Zwischenformen (Bindeglieder oder connecting links), als sie auf der Stammlinie der Tetrapoden liegen und dokumentieren, in welcher Reihenfolge sich die Tetrapoden-Merkmale evolutionsgeschichtlich entwickelt haben. Schema nach DAESCHLER et al. (2006, S. 761f.).
Die Kritik der Evolutionsgegner am Status von Tiktaalik
Kurz nach der Erstbeschreibung von Tiktaalik erschien auf der kreationistischen Homepage Genesisnet eine interessante Stellungnahme von Reinhard JUNKER. Dieser stellt zunächst fest, dass "Tiktaalik … ein Merkmalsmosaik [besitzt], das gut in einen Übergangsbereich zwischen Fischen und Vierbeinern passt - hier geht ein Punkt an die Evolutionstheorie" (JUNKER 2006). Diese Feststellung ist im Grundsatz erfreulich, da sie sich positiv von den Auslassungen anderer Evolutionsgegner abhebt. Allerdings stellt JUNKER auch kritische Bemerkungen an, die sachlich fragwürdig sind. Er schreibt beispielsweise:
"Dass Merkmale von Tiktaalik besonders hervorgehoben werden, die als 'Übergangsmerkmale' gedeutet werden können, ist legitim. Dennoch zeigt die Brustflosse insgesamt doch eher einen fischartigen Charakter (Ahlberg & Clack 2006, 748). Ein Vergleich mit anderen Formen aus dem Übergangsbereich Fische - Vierbeiner macht dies deutlich."
Abgesehen von der problematischen Differenzierung "fischartig" contra "tetrapodenartig": Entscheidend ist aus evolutionärer Sicht, dass die Brustflossen von Tiktaalik fortschrittlicher waren als die von Panderichthys, aber ursprünglicher als die von Ichthyostega und Acanthostega. Dies wird in Abb. 3 ersichtlich. AHLBERG & CLACK (2006, S. 748) bemerken hierzu:
"Panderichthys showed us a morphology that could be interpreted as directly intermediate between osteolepiform and tetrapod. But only the similar yet 'upgraded' morphology in Tiktaalik demonstrates that this interpretation is correct: this really is what our ancestors looked like when they began to leave the water."
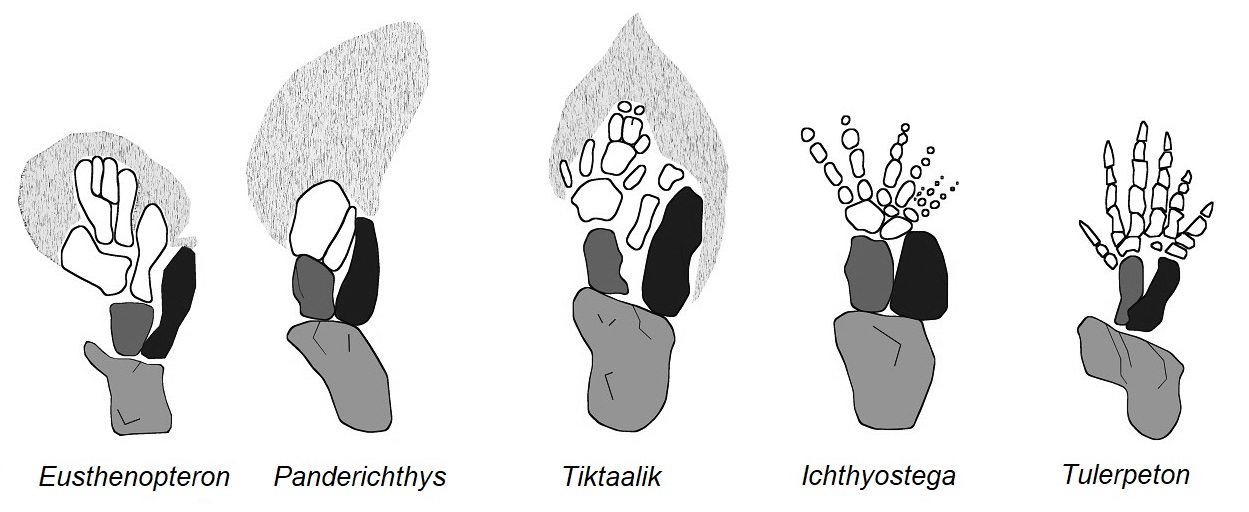
Abb. 3 Skelettbau der Vorderextremitäten verschiedener Stammtetrapoden. Die Gliederung der Brustflosse von Tiktaalik vermittelt in zahlreichen Einzelmerkmalen. Erste Fingerknochen deuten sich an, wenngleich diese noch in Flossenstrahlen enden. Umgezeichnet nach SHUBIN et al. (2006)
Fischartige oder tetrapodenartige statt intermediäre Fossilien?
JUNKER bemerkt ferner unter Bezugnahme auf PENNISI:
"Pennisi (2006) stellt in ihrem Kommentar in Science fest, dass die bislang bekannten Fossilien entweder vornehmlich fischartig oder tetrapodenartig waren statt wirklich intermediär zu sein. Offenbar erlaubt erst die verbesserte Datenlage, den bisherigen Stand des Wissens weniger geschönt darzustellen."
Genau genommen ist die Forderung nach intermediären Fossilien zwischen "eher fischartigen" und "tetrapodenartigen" Organismen unsinnig. Zum einen ist die Tetrapoden-Gruppe in die Gruppe der Fleischflosser eingeschachtelt. Das heißt, es existiert keine geschlossene Abstammungsgemeinschaft der "Fische" (PATTERSON 1982). Es existieren nur fein verästelte Verzweigungen innerhalb des Fleischflosser-Taxons (vgl. Abb. 2), zu dem auch die Tetrapoden zählen. So sind die Lungenfische und Quastenflosser mit den Vierbeinern näher verwandt als beispielsweise mit den Knorpelfischen oder Strahlenflossern. Die Beschreibung "fischartig" contra "tetrapodenartig" korreliert also mit keiner natürlichen Klassifikation und entspricht in diesem Sinn keiner wissenschaftlichen Kategorie (TAYLOR 2021, S. 3380).
Zum anderen suggeriert die Aussage, die bekannten Fossilien seien nicht "intermediär", dass aus evolutionärer Sicht der gesamte Organismus eine harmonische, kontinuierliche Veränderung seiner Merkmale zu bezeugen habe. Eine solche Evolution gibt es nicht! Intermediär können verschiedene Merkmale sein, nicht ein Organismus als Ganzes. Aufgrund funktioneller oder entwicklungsgenetischer Zwänge (Bürden) werden immer diverse Merkmale in ihrer Entwicklung zurückbleiben, während andere evolvieren.
Artaufspaltungen gestalten das Bild noch komplizierter: Wenn Populationen unterschiedliche Nischen bevölkern, wirken auf die gleichen Merkmale oft verschiedene Selektionsdrücke. Als Folge dessen werden sich unterschiedliche Merkmale sowohl innerhalb eines Organismus als auch zwischen verschiedenen Arten mit unterschiedlicher Geschwindigkeit entwickeln (CARROLL 1997). Das Ergebnis ist weit mehr eine Mosaik-Evolution mit diskontinuierlichen, teils inkongruenten Merkmalsveränderungen (vgl. beispielsweise MAHNER 1986; FUTUYMA 1990; RIEDL 2003; FELICE & GOSWAMI 2018; NEUKAMM & BEYER 2023).
Aufgrund der bekannten Prozesse der Artbildung und Artdifferenzierung weist Tiktaalik, wie jedes connecting link, ursprüngliche und abgeleitete Merkmale nebeneinander auf statt in seiner Gesamtheit intermediär zu sein.
Mit jeder Übergangsform werden weitere gefordert
"Daeschler et al. (2006, 757) [meinen], dass ... der Ursprung der wichtigen Tetrapodenmerkmale jedoch in Dunkeln verblieben sei."
Diese Aussage ist richtig, ein Argument gegen Evolution folgt daraus nicht.
JUNKER & SCHERER (1998, S. 217) merkten zur Evolution der Tetrapoden noch an, die "Homologisierung zwischen der viergliedrigen Tetrapoden-Extremität und Flossenknochen von Fischen" sei aufgrund des Fehlens geeigneter Übergangsformen "ungeklärt". Sie betonen, die Urtetrapoden seien somit nicht von tetrapodenähnlichen Fischen ableitbar. Mit Tiktaalik ändert sich die Situation grundlegend.
Es stellt sich die Frage, wie viele Zwischenformen es noch braucht, damit sie die Evolutionsgegner endlich als Belege für die Darwin'sche Abstammungstheorie akzeptieren. Da JUNKER einräumt, mit dem Fund sei die morphologische Lücke verkleinert worden, wundert man sich, weshalb er wieder nach dem gleichen Argumentationsschema verfährt, das lautet:
"Präsentiert man eine Zwischenform, so wollen die Kreationisten zwischen dieser und der Ahnenform eine weitere Zwischenform und so fort ... dies lässt sich ad infinitum weitertreiben." (MAHNER 1986, S. 62)
Wie erwähnt halten wir JUNKER zugute, dass er einräumt, das Merkmalsmosaik von Tiktaalik passe gut in einen Übergangsbereich "zwischen Fischen und Vierbeinern". Aber angesichts der folgenden Aussage ist diese halbherzige Konzession ein "vergiftetes Geschenk" an die Evolutionsbiologie:
"Die Unterschiede zwischen Tiktaalik und gefingerten Gattungen wie Acanthostega sind erheblich … Ahlberg & Clack (2006, 748) weisen darauf hin, dass der Erwerb von Fingern, von Tiktaalik ausgehend, eine erhebliche Umorganisation ('developmental repatterning') erfordern würde. Der achtfingrige oberdevonische Tetrapode Acanthostega … war höchstwahrscheinlich ausschließlich wasserlebend; seine Extremitäten waren relativ unbeweglich. Insgesamt eignet sich diese Gattung daher nicht als vermittelnde Form zwischen Tiktaalik und landlebenden Tetrapoden."
"Daher"? Diese Aussage könnte den Eindruck erwecken, als würde JUNKER hier eine Schlussfolgerung von AHLBERG & CLACK wiedergeben. Die Spezialisten für frühe Tetrapoden gelangen im Rahmen ihrer Merkmalsanalysen zu genau dem entgegengesetzten Schluss und sprechen diesbezüglich von einem "firm step from water to land". Die von den Autoren publizierten Kladogramme (AHLBERG & CLACK 2006, S. 747; DAESCHLER et al. 2006, S. 761) stehen im Widerspruch zu JUNKERs Einschätzung.
Die Evolution fährt auf vielen Gleisen gleichzeitig
SHUBIN et al. (2006, S. 768) weisen darauf hin, dass der primitivere Rhizodontier Sauripterus mit Tiktaalik ein fortschrittliches Merkmal teilte, das nicht auf gemeinsame Abstammung zurück zu führen sei: die Fingerknochen. Vielmehr sei dieses Merkmal in beiden Gruppen konvergent, das heißt unabhängig voneinander entstanden. Der insgesamt fortschrittlichere Acanthostega wiederum behielt einige Primitivmerkmale bei, die bei dem ursprünglicheren Tiktaalik fortschrittlicher waren.
Diese sogenannten Inkongruenzen kommentiert JUNKER wie folgt:
"Tiktaalik hatte deutlich andere 'Übergangsmerkmale' auf dem Weg zum Landleben als Acanthostega. Der Weg aufs Land konnte nicht über beide Formen zugleich führen, es sei denn, er wurde mindestens zweimal unabhängig durchlaufen, womit dann aber die Vierbeinigkeit nicht mehr als Schlüsselmerkmal (d. h. als Hinweis auf gemeinsame Vorfahren) gelten könnte, sondern konvergent entstanden wäre."
JUNKER wiederholt hier eine Erkenntnis, die mit unserem Wissen über Artbildungsprozesse seit einem halben Jahrhundert konformgeht: Wie erwähnt begünstigen Artaufspaltungen eine Mosaik-Evolution mit individuell verschiedenen Evolutionsdynamiken in den voneinander isolierten Entwicklungslinien (vgl. MAYR 1967, S. 465f.; MACFADDEN 2005). Gelegentlich evolvieren die gleichen Merkmale in verschiedenen Populationen sogar in unterschiedliche Richtungen (FUTUYMA 2015, S. 35). Diese "mehrgleisig" ablaufende (nicht-lineare) Evolution erzeugt in der Gesamtschau das Bild einer disharmonischen, teils inkongruenten Merkmalsverteilung zwischen den verschiedenen Linien.
Entgegen JUNKER bedeutet dies nicht, dass die Vierbeinigkeit nicht mehr als ein von einem gemeinsamen Vorfahren der Tetrapoden erworbenes Merkmal gelten könne. Die von SHUBIN et al. (2006, a.a.O.) diskutierten Konvergenzen bedeuten nur, dass sich die Extremitäten in einigen Entwicklungslinien unabhängig voneinander ähnlich spezialisiert haben.
Dass die von JUNKER kritisierte Sicht der Evolution keine zeitgemäße ist, belegt auch der folgende Einwand:
"Schon länger ist klar, dass auch das berühmte Ichthyostega … vor noch nicht langer Zeit die Ikone für den Übergang vom Wasser- zum Landleben, deutlich von einer vermittelnden Position entfernt ist … Nicht nur der Bau der Flossen, sondern auch der Schädelbau passt insgesamt nicht in eine evolutive Reihe von tetrapodenähnlichen Fischen hin zu frühen Tetrapoden. Auch in dieser Hinsicht würde sich Acanthostega auf einer anderen 'Schiene' bewegen, wenn man die relevanten fossilen Gattungen in evolutionäre Linien einfügen wollte".
Kein phylogenetischer Systematiker hat je gefordert, dass Panderichthys, Tiktaalik, Acanthostega und Ichthyostega auf "einer Schiene" von "tetrapodenähnlichen Fischen zu den frühen Tetrapoden" führen müssten. Im Gegenteil, die phylogenetische Systematik beschäftigt sich mit Artaufspaltungen, das heißt mit Verzweigungsmustern. Und jede Verzweigung führt zwangsläufig zu zwei unterschiedlichen Entwicklungs-"Schienen".
Wo will man in dem evolutionären Dickicht von Verzweigungen, denen entlang sich die Stammesgeschichte ihre gewundenen Wege bahnt, überhaupt so etwas wie eine "Hauptschiene" der Entwicklung ausmachen? Der kladogenetische Aspekt der Evolution lässt keine "evolutiven Reihen" oder unilineare Übergangsserien zu, sondern erzeugt komplexe Übergangsfelder. Arten, die aufgrund ökologischer Sonderung (bzw. selektionsbedingt) auf "unterschiedlichen Schienen" evolvieren, treiben die Entwicklung teils unterschiedlicher Merkmale voran, wogegen bei anderen Arten diverse Merkmale konvergent entstehen.
Die Evolution lässt sich jeweils nur über gewundene Wege bis zu einem letzten gemeinsamen Vorfahren zurückverfolgen,
"aber kein Weg verläuft gerade, und alle führen über seitliche Schritte von einem Artbildungsereignis zum nächsten in die Vergangenheit, nicht über eine Abstammungsleiter der kontinuierlichen Veränderung" (GOULD 2002, S. 92).
Wenn JUNKER daraus folgert, dass die Gattung Acanthostega "nicht als vermittelnde Form zwischen Tiktaalik und landlebenden Tetrapoden infrage" komme, zieht er einen falschen Schluss, denn er zeichnet die Evolution einfacher, als sie aufgrund des aktuellen Hintergrundwissens angenommen werden muss.
Resümee
Im Einklang mit JUNKER lässt sich festhalten, "dass Tiktaalik das Spektrum von Fischen mit tetrapodenartigen Merkmalen erweitert und in diesem Sinne … einen Baustein für evolutionäre Übergangshypothesen darstellt. Die morphologische Lücke zwischen manchen Formen wird mit dem neuen Fund verkleinert." Des Weiteren zeigt sich hier erneut, was "Makroevolution" ausmacht: Von den älteren Fossilschichten aus betrachtet nähern sich die Arten in stufenweiser Abänderung den heutigen Formen an. Wenn Mosaikformen wie Eusthenopteron, Panderichthys, Acanthostega, Ichthyostega und Tiktaalik keine schöne Bestätigung der Darwin'schen Abstammungstheorie sind, was dann?
Literatur
AHLBERG, P. E. & CLACK, J. A. (2006) A firm step from water to land. Nature 440, S. 747-749.
CARROLL R. L. (1997) Patterns and processes of vertebrate evolution. Cambridge University Press, Cambridge.
DAESCHLER, E. B.; SHUBIN, N. H. & JENKINS, F. A. (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, S. 757-763.
DALTON, R. (2006) The fish that crawled out of the water. A newly found fossil links fish to land-lubbers.
FELICE, R. N. & GOSWAMI, A. (2018) Developmental origins of mosaic evolution in the avian cranium. PNAS 115, S. 555–560.
FUTUYMA, D. J. (1990) Entwicklungsintegration und Makroevolution. In: ders.: Evolutionsbiologie. Birkhäuser, Basel, S. 497-498.
FUTUYMA, D. J. (2015) Can modern evolutionary theory explain macroevolution? In: SERRELLI, E. & GONTIER, N. (Hg.) Macroevolution. Explanation, interpretation, and evidence (S. 29–85). Springer-Verlag, Heidelberg.
GOULD, S. J. (2002) Illusion Fortschritt. Die vielfältigen Wege der Evolution. Fischer-Verlag, Frankfurt.
JUNKER, R. (2006) Tiktaalik - ein erstklassiges Bindeglied?
JUNKER, R. & SCHERER, S. (1998) Evolution. Ein kritisches Lehrbuch. Weyel-Verlag, Gießen.
MACFADDEN, B. J. (2005) Fossil horses – evidence for evolution. Science 307, S. 1728–1730.
MAHNER, M. (1986) Kreationismus - Inhalt und Struktur antievolutionistischer Argumentation. Pädagogisches Zentrum, Berlin.
MAYR, E. (1967) Artbegriff und Evolution. Verlag Paul Parey, Singhofen.
NEUKAMM, M. & BEYER, A. (2023) Woher wissen wir, dass Vögel lebende Dinosaurier sind?
PATTERSON, C. (1982) Cladistics and classification. New Scientist 94, S. 303-306.
RIEDL, R. (2003) Riedls Kulturgeschichte der Evolutionstheorie. Springer-Verlag, Berlin.
SHUBIN, N. H.; DAESCHLER, E. B. & JENKINS, F. A. (2006) The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, S. 764-771.
TAYLOR, H. (2021) Whales, fish and Alaskan bears: interest-relative taxonomy and kind pluralism in biology. Synthese 198, S. 3369–3387.
Autor: Martin Neukamm
