![]()
PDF-Version
Buchbesprechung
Das Argument der Designfehler (unintelligentes Design)
Warum sich der Eindruck von Schöpfung in der Evolution verflüchtigt

Ein Argument gegen den "intelligenten" Ursprung biologischer Merkmale nimmt auf die Existenz sinnloser Strukturen, Umwege und funktioneller Mängel Bezug, die teils mit einer Herabsetzung der Fitness ihrer Besitzer einhergehen (kurz: Design- oder Konstruktionsfehler). Solche Strukturen sind genau das, was man aus Sicht einer Evolution erwartet, die nicht vorausschauend planen, sich ihres stammesgeschichtlichen Erbes nicht entledigen und Organismen nicht "wegen Umbaus vorübergehend außer Betrieb nehmen" kann. Das unterscheidet "ungelenkte" Naturprozesse von zielorientierter Planung, die nicht notwendigerweise an diese Bedingungen gebunden ist.
Da verschiedene "Design-Indizien" auf die "Zweckmäßigkeit bzw. Zielorientierung" biotischer Strukturen abheben (JUNKER & WIDENMEYER 2021, S. 201), wird der Prima-facie-Eindruck von Planung umso fragwürdiger, je deutlicher und je häufiger unzweckmäßige, dysfunktionale oder nicht zielorientierte Strukturen und Umwege (Design- oder Konstruktionsfehler) scheinbar "hochgradig speziell ausgeführte" Merkmale überlagern. Es gilt:
"Während optimale Passung auch als Beleg für teleologische Ansätze … gewertet werden kann, können suboptimale Performance, Konstruktionsdefekte … und das verbreitete Vorkommen von Junk-DNA nur im Lichte einer Theorie erklärt werden, in der Begriffe wie 'historischer Zufall', 'Variation' und 'Selektion' vorkommen." (MAHNER & BUNGE 2000, S. 342)
Das vorliegende Buch von JUNKER & WIDENMEYER versucht in zwei Kapiteln (S. 212f. sowie S. 275–287) dieses Argument zu entkräften. Bevor wir uns den Einwänden widmen, sollen zunächst klassische Konstruktionsdefizite angeführt und daran exemplarisch die evolutionstheoretischen Erklärungen erläutert werden.
Klassische Beispiele für Konstruktionsfehler
Mit Blick auf die funktionale Komplexität und "Genialität lebender Konstruktionen" (siehe S. 259) ist es bei Evolutionsgegnern Gang und Gäbe, auf die Konstruktionsabsicht eines intelligenten Planers zu schließen. Doch bereits HASS (1979) zeigte am Beispiel des Urogenitalsystems der Vertebraten, dass sich die Konstruktionswidrigkeiten mit dem Adjektiv "genial" nur schwer charakterisieren lassen (nach MAHNER 1986, S. 76):
Die Embryonalentwicklung schlägt eine Reihe von Umwegen ein mit dem Ergebnis, dass die Eizellen einen umständlichen Weg vom Ovar zum Ostium tubae und Ovidukt zurückzulegen haben, der sich angesichts des Risikos von Eileiter- und Bauchhöhlenschwangerschaften als potentiell lebensgefährlich herausstellt. Müsste man von einem genialen Konstrukteur nicht geradezu erwarten, dass solche Fehler schon während der Planung entdeckt und gar nicht erst begangen werden? Wozu wurde kein eigener Ausführgang für die Keimdrüsen angelegt und dafür auf Umwege wie Kloakenbildung verzichtet?
Auf derlei Konstruktionsmerkwürdigkeiten stößt man, wie RIEDL (1984, S. 18) bemerkt, allein bei der "Krone der Schöpfung" dutzendfach:
"Das Heer der konstitutionellen Krankheiten ist die Folge: Schwindel, Bandscheiben- Schwäche, Leistenbruch, Hämorrhoiden, Krampfadern, Plattfüße. Ja, vieles ist überhaupt unreparierbar verbaut."
Ein eklatanter Fall von Dysfunktionalität ist die Immunpathogenese. Dabei handelt es sich um eine fatale Überreaktion des Immunsystems, die Betroffene regelmäßig umbringt. Sie beginnt mit einer unkontrollierten Produktion von Immun-Botenstoffen und führt unbehandelt zum Organversagen. Dieser Effekt ist oftmals für die tödlichen Verläufe von CoViD-19, Ebola und Influenza verantwortlich oder wesentlich daran beteiligt.
Ein weiteres Beispiel ist unsere Netzhaut. Sie ist bei allen Wirbeltieren verkehrt herum ("invers") in den Augenbecher eingebaut (Abb. 1). Absurderweise passiert das Licht ein Geflecht von Blutgefäßen und Nervenzellen, bevor es auf die Sehzellen trifft, und an der Austrittsstelle des Sehnervs entsteht ein "blinder Fleck" auf der Netzhaut.
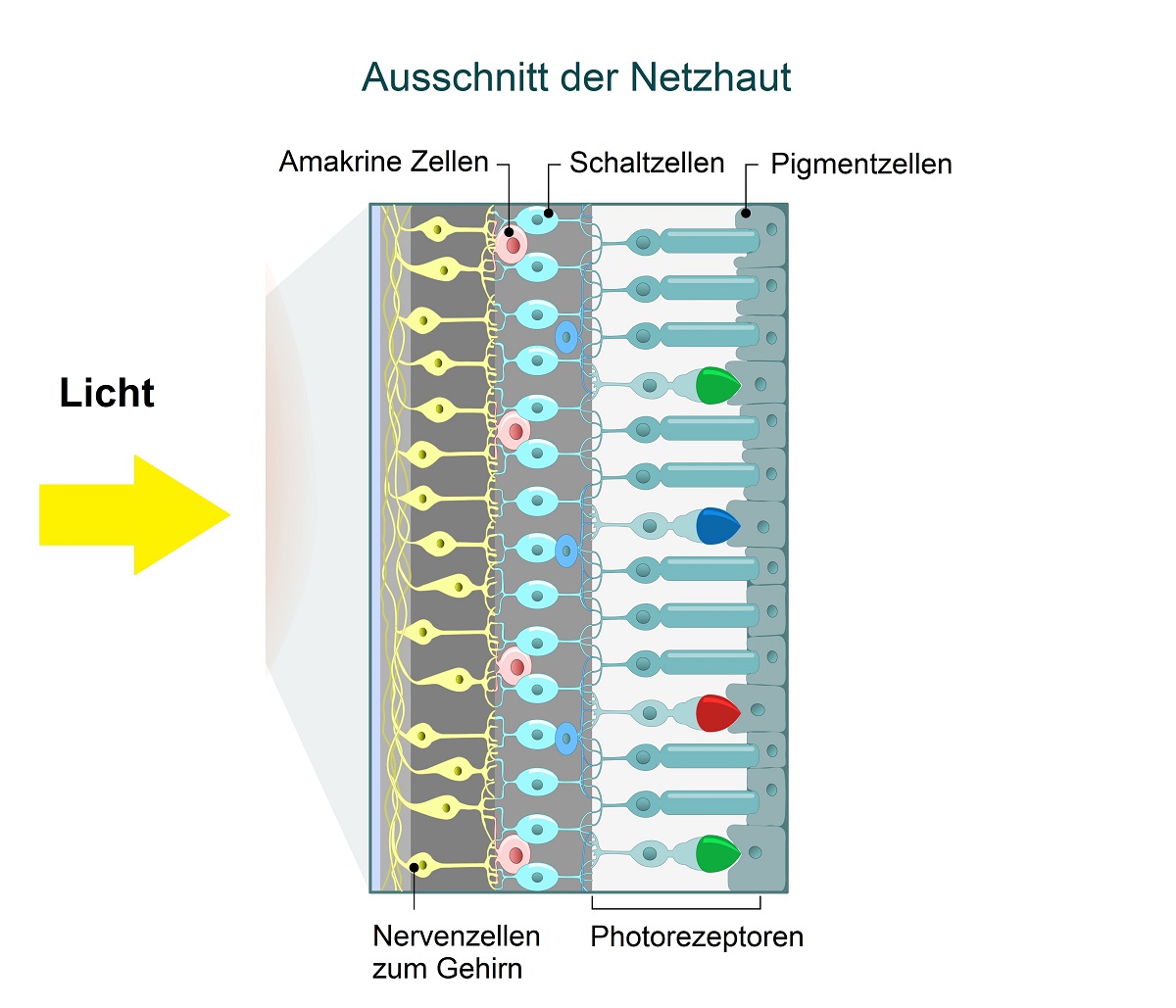
Abb. 1 Inverser Bau der Netzhaut: Die Sehzellen (Photorezeptoren) befinden sich auf der dem Licht abgewandten Seite, während die Nervenzellen dem Licht zugewandt sind. Das ist kein cleverer Schachzug: Das Licht muss ein Netz aus Blutgefäßen und Nervenzellen passieren, bevor es die Sehzellen erreicht. Dies geht mit Lichtverlust, optischen Verzerrungen und Einbußen der Sehschärfe einher. Zudem entsteht an der Austrittsstelle des Sehnervs ein "blinder Fleck".
Nicht minder kurios ist der Verlauf des rückläufigen Kehlkopfnervs (Stimmnerv) der Säugetiere (Nervus laryngeus recurrens). Er zweigt vom zehnten Hirnnerv (Nervus vagus) ab, führt aber nicht direkt zum Kehlkopf, wie man es unter dem Aspekt eines zielgerichteten Ursprungs erwarten würde, sondern wenige Millimeter daran vorbei! Unterhalb des Halses taucht der Nerv unerwartet im Brustkorb ab und vollführt in der Nähe des Herzens eine sinnlose Schleife um den Aortenbogen und die Unterschlüsselbeinarterie ("Rekurrens-Schlinge"). Schließlich legt er zwischen Luft- und Speiseröhre den ganzen Weg zurück zum Hals, wo er endlich an seinen Bestimmungsort, dem Kehlkopf, ankommt. Dort innerviert sein Endast den unteren Kehlkopfnerv, die Schleimhaut unterhalb der Stimmlippen sowie die Kehlkopfmuskulatur (STEINBRÜCK et al. 2008). Beim Menschen beträgt der Umweg nur etwa 15 cm und ist damit noch recht kurz. Bei der Giraffe aber beläuft sich der Umweg auf unglaubliche 4,5 Meter!
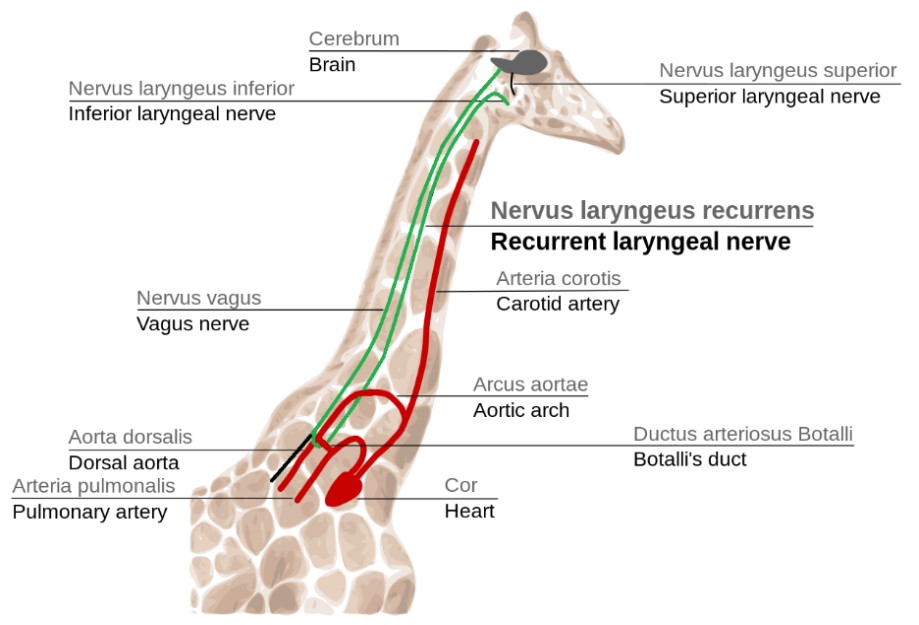
Abb. 2 Rekurrensschlinge bei der Giraffe, dargestellt in grün. Bildquelle: Dr Bug (Vladimir V. Medeyko) in some jurisdictions may be treated as defivative of Oakland_Zoo_dsc_2658.jpg: Eugenia and Julian from San Jose - Bay Area, California, USA, GiraffaRecurrRu, Rekurrensschlinge grün eingefärbt, CC BY-SA 2.0.
{kind=link}
{kind=link}
Ein letztes Beispiel: Der Brutbeutel der Koalas ist nicht, wie bei den meisten Beuteltieren, nach oben hin offen, sondern nach unten. Unter australischen Rangern kursiert daher der makabre Witz: "Der Koala fällt nicht weit vom Stamm und stirbt da auch" (STEFFENS 2007, S. 17). Zum Glück besitzen Koala-Babys einen ausgeprägten Klammerreflex, der solches zumeist verhindert. All diese Strukturen ergeben aus teleologischer Perspektive keinen Sinn, vergeuden unnötig Ressourcen, sind suboptimal oder gar lebensgefährlich. Inwieweit aber kann die Evolutionstheorie ihre Herkunft erklären?
Die evolutionstheoretische Erklärung: drei Beispiele
Die Rekurrens-Schlinge des rückwärtigen Kehlkopfnervs erklärt sich zwanglos, wenn wir zugrunde legen, dass die Evolution nicht zielgerichtet verlief, während unsere Vorfahren Kiemenatmer waren: Bei den "Fischen" schlägt der Nerv nämlich keinen Umweg ein, sondern führt als sechster Kiemenbogennerv direkt zum Schlundbogen. Waren die Kiemen und Aortenbögen mit ihrem Geflecht an Blutgefäßen und Nerven erst einmal im Bauplan der "Fische" etabliert, musste jede evolutionäre Weiterentwicklung die Funktionsfähigkeit des Merkmalsgefüges gewährleisten.
Als die Kiemen in der Evolution verschwanden und die Landwirbeltiere Hals und Kehlkopf entwickelten, war es entwicklungsbiologisch nur noch möglich, die Aortenbögen zu modifizieren. Es war nicht möglich, sie außer Dienst zu stellen und das Gewirr aus Blutbahnen, Organen und Nervensträngen grundlegend neu zu ordnen. Letzteres würde verhindert haben, dass beim Verlängern des Halses und beim Abstieg (Deszensus) des Herzens in den Brustkorb der vierte bis sechsten Aortenbogen mit nach unten wandert und den Nervus laryngeus recurrens mitnimmt. Doch Mutationen, die das Chaos im Brustkorb beseitigen, sind faktisch unmöglich, weil sie viele Merkmale gleichzeitig ändern müssten. Somit ist aus selektionstheoretischer Perspektive der jeweilige Nutzen einer sukzessiven Verlängerung des Nervs höher als eine Neugestaltung des Bauplans.1)
Analoges gilt für die Netzhaut: Beim plattenförmigen Nervensystem an der Körperoberfläche einfacher Deuterostomier zeigen die Sinneszellen zweckmäßigerweise zum Licht. Durch Einrollen ins Körperinnere entsteht aus der Neuralplatte das Neuralrohr als erste embryonale Entwicklungsstufe des zentralen Nervensystems höherer Tiere. Dies bewirkt ein "Umkrempeln" des Gewebes, sodass die lichtempfindlichen, primären Sinneszellen im Innern des Neuralrohrs liegen (FRANZE & GROSCHE 2008, S. 701).
Das Neuralrohr tritt bereits bei augenlosen Chordatieren auf, den Vorfahren der Wirbeltiere. Bei den Wirbeltieren bilden sich aus dem Neuralrohr später Gehirn, Rückenmark und Augen. Unter der Herrschaft des PAX-6-Mastergens quellen aus dem hinteren Bereich des Vorderhirns Wülste hervor, die sich zum Augenbecher umformen (MÜLLER & HASSEL 2018, S. 475). Aufgrund dieses Entwicklungszwangs (engl.: developmental constraint) kommt es zur Inversion der Netzhaut (Abb. 3).
Jene embryogenetische Weichenstellung, die die Inversion der Netzhaut herbeiführt, trafen also schon die Vorfahren der Chordatiere. Sie geht auf eine Zeit zurück, bevor sich die Entstehung von Wirbeltieraugen abzeichnete. Diese "Vorentscheidungen" ließen später keine Alternativen mehr zu. Denn ihr genetisches Erbe zu modifizieren, bis die Sehzellen "richtig" im Augenbecher liegen, würde es erfordern, die embryonale Anlage des Nervensystems von Grund auf umzugestalten. Für Prozesse, die keinen Plan kennen, ist das nicht leistbar, also blieb die Differenzierung von Stammzellen des Neuralrohrs in ihrer besonderen topographischen Lage zu den Sinneszellen, wie sie war.
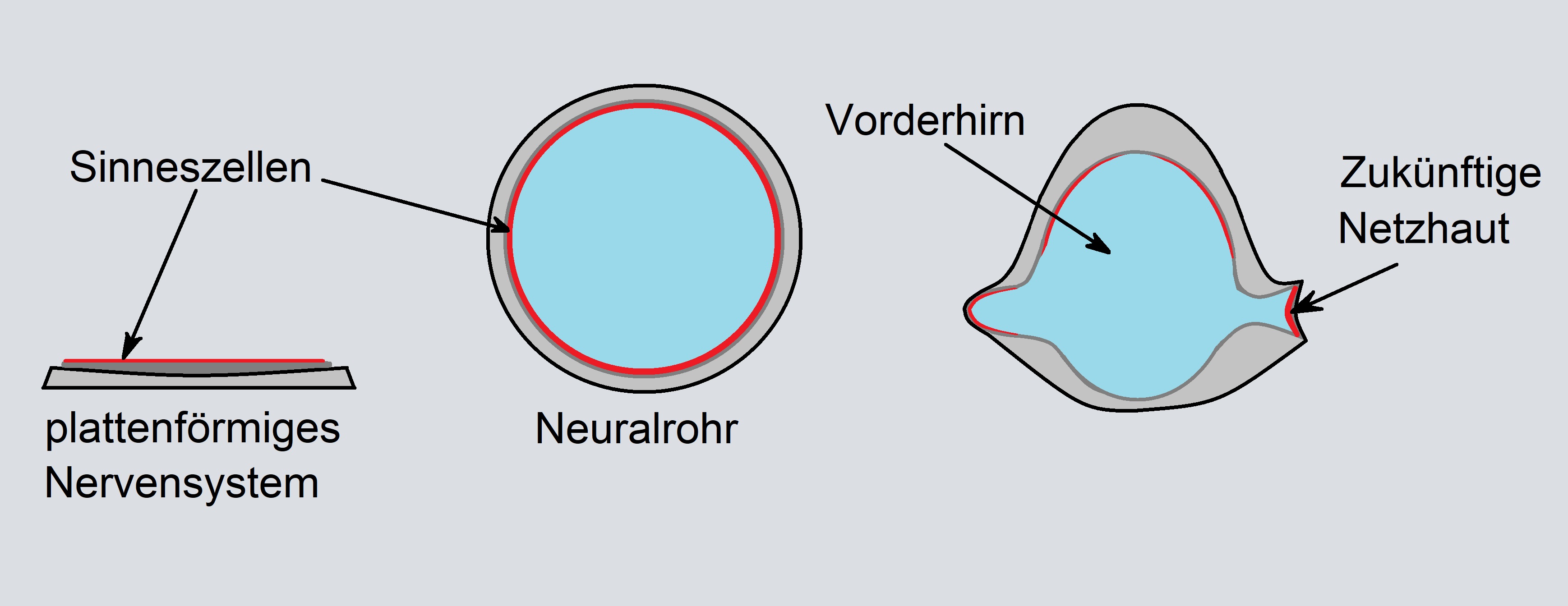
Abb. 3 Entwicklung der inversen Netzhaut, ausgehend vom platten Nervensystem einfacher Deuterostomier (z. B. Seesterne) zum ins Körperinnere eingesenkten Neuralrohr als Anlage des Gehirns und Rückenmarks. Gezeichnet nach FRANZE & GROSCHE (2008, S. 701).
Dagegen lässt sich nicht sinnvoll erklären, warum sich ein Designer Entwicklungszwängen unterwarf, die ihren Ursprung in historisch älteren Bauplänen haben. Er soll doch nach Ansicht von Intelligent Design (ID) in der Lage sein, überlegt zu handeln, die Merkmale planvoll, zielgerichtet und "frei" zu kombinieren.
Je stärker die biologischen Merkmale durch Konstruktionsfehler überlagert sind, desto weniger genügen sie Zweck-Mittel-Beziehungen und Funktionsanforderungen – und desto mehr verliert sich der Eindruck von Planung. Sie passen hervorragend zu evolutiven Prozessen, die kein Ziel kennen.
Die Entwicklungsbiologen MÜLLER & HASSEL (2018, S. 627) resümieren:
"Die (nahezu) perfekte Konstruktion bestimmter Augen verleitet manche Menschen zu der Annahme, dass ein intelligentes Wesen diese Augen geschaffen haben müsse, natürliche Evolution könne sie nicht hervorgebracht haben. Zunehmendes Wissen um Homologie auf molekularem Niveau, um Zwischenstufen in der Konstruktion wie auch um suboptimale Lösungen (z. B. inverses Auge der Wirbeltiere, schlechte optische Qualität des dioptrischen Apparates) machen Annahmen über 'intelligentes Design' aber keineswegs zwingend. Im Gegenteil unterstützen die Daten … die Sicht, dass bekannte Mechanismen der Evolution letztlich eine Vielfalt brauchbarer Lösungen hervorbrachten."
Der Grund für die verkehrte Öffnung des Koala-Brutbeutels ist ebenfalls schnell erklärt: Koalas haben wombatartige Vorfahren, und Wombats graben mit Vorliebe in der Erde: Sie befördern das Erdreich mit ihren schaufelartigen Pfoten nach hinten wie ein Bagger. Wäre der Beutel des Vorfahren nach vorne offen gewesen, wären die Jungen mit Dreck zugeschüttet worden. Also zeigte die Öffnung nach hinten, und als die Nachfahren dieser Tiere auf Bäume kletterten, nahmen sie diese "Konstruktion" mit; es war zu kompliziert, sie zu ändern (WILLIAMS 2006, S. 63).
Einwände gegen das Argument der Designfehler und Gegenargumente
A. Das Argument sei "theologisch" kontaminiert und somit unzulässig
JUNKER & WIDENMEYER (2021) entgegnen, die evolutionäre Argumentation setze stillschweigend bestimmte theologische Annahmen voraus. Sie sei nur zielführend, wenn angenommen werde, dass ein Schöpfer perfekte Merkmale hätte hervorbringen müssen, was ID aber gar nicht behaupte:
"[S]olche Argumente [enthalten] bestimmte (theologische) Annahmen über einen hypothetischen Schöpfer, z. B. dass ein möglicher Schöpfer gute, wenn nicht zwingende Gründe gehabt haben müsste, perfekte Strukturen (in unserem Sinne) hervorzubringen. Diese meist stillschweigend gemachten Annahmen werden meistens nicht einmal versucht zu begründen …" (S. 212)
Doch das Argument gegen "Design" setzt keineswegs voraus, dass intelligente Planung perfekte Ergebnisse haben müsse. Es setzt lediglich voraus, dass simple Fehler und vorhersehbare Schwierigkeiten vermieden werden – so wie es jede intelligente Planung, auch weit weg von der Perfektion, leisten würde. Anders gesagt: "Schlechtes" Design passt besser zu nicht-zielgerichteten Prozessen (Evolution) als zu zielgerichtetem, überlegtem Handeln (KOJONEN 2016, S. 152-153).2) Entsprechend räumt auch RAMMERSTORFER (2006) ein, dass "banale Fehler" den Design-Ansatz schwächen würden, denn:
"Stümperhaftes Design passt insgesamt (wie oben besprochen) hervorragend zu Prozessen, die keinen Plan kennen. Damit würde auf dieser Ebene ('Design-Fehler') ein gutes Argument für Evolution und gegen Planung existieren." (S. 90)
"Ist das [gute Design] wirklich zu erwarten, wenn man als Ursprungserklärung einen ungelenkten Prozess postuliert...? Oder wäre in diesem Fall nicht genau das Gegenteil zu erwarten: Überwiegend 'schlechte Designs' (maximal optimiert vielleicht, aber trotzdem zweitklassig) und hin und wieder gute Designs?" (S. 88)
Das Argument der Designfehler hat also nichts mit Theologie zu tun; es orientiert sich schlicht an den ontologischen und methodologischen Leitsätzen von ID, wie: "Planmäßigkeit lässt einen Planer vermuten" oder: "intelligente Konstruktionen legen eine intelligente Entstehungsursache nahe". Oder wie RAMMERSTORFER (2003) bemerkt:
"Der Schluss von Planmäßigkeit, Zielgerichtetheit, wie sie in der Natur überall zu finden ist, auf einen Planer, ist ganz einfach der naheliegende Schluss."
Es ist daher völlig legitim, neben den zahllosen Schilderungen (gewiss!) faszinierender und komplexer Bau- und Funktionszusammenhänge in der Natur auch auf die eklatanten Konstruktionsdefizite hinzuweisen und kritisch zu hinterfragen, ob sich Ingenieuren, Technikern, Morphologen und Medizinern in der Gesamtschau der Eindruck von Planmäßigkeit aufdrängte.3) Um es mit Mahner zu sagen:
"Die Planmäßigkeitsanalogie streicht den Sinn in der Natur heraus und übersieht den Unsinn. Auf gleiche oder ähnliche [= intelligente] Ursachen darf nicht geschlossen werden." (MAHNER 1986, S. 79)
Solche Überlegungen als "versteckte Theologie" zu missdeuten, ist ein durchschaubares Manöver. Schließlich bemühen die WORT-UND-WISSEN-Autoren selbst mannigfach Beispiele und Analogien mit der Technik, die den Eindruck planerischer Genialität und Optimalität vermitteln sollen. Steht die Analogie dem Planmäßigkeits-Argument jedoch im Weg, wird sie geleugnet. Dieser argumentative Doppel-Standard ist ein Kennzeichen von Kritikimmunisierung.
Im Übrigen ist der Einwand schon deshalb obsolet, da die Autoren selbst, wie überhaupt die meisten ID-Vertreter, den Schöpfergott als superintelligenten, perfekten Konstrukteur voraussetzen. Das ist keineswegs das traditionelle biblische Gottesbild, sondern entstand erst im 18. und 19. Jahrhundert in der Auseinandersetzung mit der Naturwissenschaft. Nicht nur die offenkundige Planlosigkeit der Evolution, sondern auch die Grausamkeit der Natur und das Aussterben von über 99 Prozent der Arten, die jemals existierten, passen nicht zu diesem Gottesbild. Aber die Diskussion um das Gottesbild sollen die Design-Advokaten mit Theologinnen und Theologen führen. In der Naturwissenschaft werden seine problematischen Konsequenzen immer nur ad hoc durch Immunisierungsstrategien wegdiskutiert, so wie dies im Folgenden (erfolglos) versucht wird.
B. Das Argument berücksichtige nicht alle Detailkenntnisse
Dieser Einwand leugnet, dass Konstruktionsfehler überhaupt existieren:
"Erstens zeigt eine Reihe von Untersuchungen, dass bei genauerer Kenntnis betreffender Organe nennenswerte Mängel nicht nachweisbar sind. Dem Argument der Konstruktionsfehler liegt oft nur ein Mangel an Kenntnissen über den betreffenden Gegenstand zugrunde - und, wie es scheint, weltanschaulich motivierte Vorurteile." (S. 212)
"Weitet man … die Perspektive und betrachtet zusätzlich zur spezifischen Struktur der Netzhaut des jeweiligen Organismus deren anatomische und physiologische Umgebung, verliert das Argument der Fehlkonstruktion an Gewicht." (S. 280)
Bezüglich der inversen Konstruktion der Netzhaut der Wirbeltiere wird auf eine Entdeckung aus dem Jahr 2007 hingewiesen, wonach es ein bestimmter Zell-Typ, die sogenannten MÜLLER-Zellen (Abb. 4) das ins Auge einfallende Licht an den streuenden Zellschichten vorbei auf die Netzhaut leiten. Diese Zellen optimieren den Sehvorgang insofern, als sie wie Lichtleiter wirken:
"Der scheinbare Widerspruch zwischen dem anatomischen und dem physiologischen Befund löste sich auf, als man intensiv weiter in die Detailforschung zur Lösung des Problems investierte." (S. 281)
Was uns hier als geniale Lösung verkauft wird, ist aber nur eine Behelfslösung, eine nachträgliche Bastelei, welche die "offensichtlich unvorteilhafte Gesamtsituation optimiert" (FRANZE et al. 2007, S. 8287; ins Deutsche MN, Hervorhebung nicht im Original). Das heißt, es wurde ein Workaround geschaffen, der die schwerwiegenden Konsequenzen des konstruktiven Mangels abmildert, ohne den Mangel selbst zu beseitigen. Das Argument der Designfehler bleibt also unangetastet; zielorientierte Lösungen offenbaren sich nicht in einer nachträglichen, behelfsmäßigen ("blinder Fleck"!) Fehlerkorrektur, sondern in vorausschauender Fehlervermeidung (MYERS 2010).
Eine weitreichende Kompensation der optischen Verzerrungen und Sehschärfe-Einbußen ermöglichen auch die MÜLLER-Zellen nicht. Dies zeigt sich an der Gefäßarmut im Bereich des schärfsten Sehens. Zahlreiche Vögel verzichten ganz auf eine vaskularisierte Netzhaut. Über ihren Fotorezeptoren winden sich keine Blutgefäße, dadurch sehen sie schärfer.
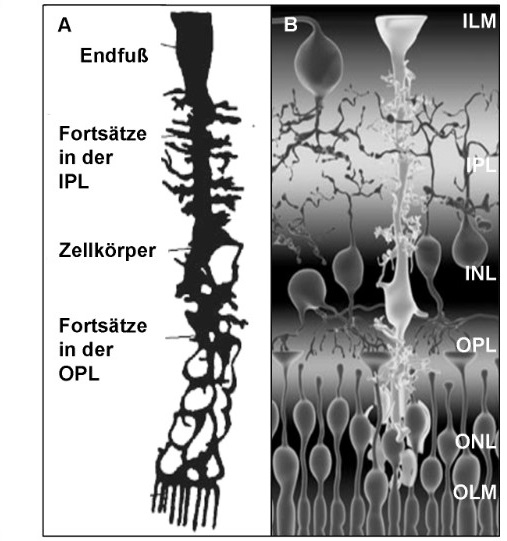
Abb. 4 Schematische Darstellung einer Müllerzelle. (A) Isolierte Zelle. (B) Zelle im retinalen Verband. Mit dem Endfuß grenzt die Müllerzelle im Bereich der inneren limitierenden Membran (ILM) an den Glaskörper. Vom Stammfortsatz in der inneren Retina zweigen Sekundärfortsätze ab und umhüllen Synapsen in der inneren plexiformen Schicht (IPL). In der inneren Körnerschicht (INL) ist der Zellkörper mit dem Zellkern lokalisiert. Sekundärfortsätze der äußeren plexiformen Schicht (OPL) stehen im Kontakt zu Synapsen der äußeren Retina. Bildquelle: EBERHARDT (2011), nach BRINGMANN et al. (2006) und LU et al. (2006).
Schlimmer noch, die Behelfslösung entpuppt sich selbst als Designfehler mit gravierenden Folgen. Die MÜLLER-Zellen haben nämlich einen erheblichen Nachteil: Sie neigen, ausgelöst durch UV-Licht, oxidativen Stress, Traumata, Infektionen, Ischämien usw., zur Hypertrophie, zur Bildung von Verdickungen und vermehrten Fortsätzen (BRINGMANN & REICHENBACH 2001). Diese sogenannte "Gliose" schädigt zunehmend Neurone bzw. Fotorezeptoren und führt unbehandelt zur Degeneration der Netzhaut bis zum völligen Sehverlust. Die physiologisch veränderten MÜLLER-Zellen sind in ein ganzes Spektrum progredienter Augenerkrankungen involviert wie Entzündungen der Augenhaut (Uveitis), Glaukomen, bis hin zu Retinopathien und Makulaödemen.
Weshalb hat der "geniale" Konstrukteur nicht die Vaskularisation samt Nährstoffversorgung und ableitenden Nerven hinter die Sehzellen verlagert und auf hypertrophierende Müllerzellen und "blinde Flecke" verzichtet? Dass es bessergeht, belegen die Kopffüßer: Da es ihr stammesgeschichtliches Erbe zuließ, sitzt die Netzhaut funktionell passend im Augenbecher. Auch die Blutversorgung ist hier eleganter gelöst (MYERS 2010).
C. Designfehler seien in Wahrheit Design-Kompromisse
Um den Eindruck von "Planung" zu retten, ist es eine beliebte Strategie, (sekundäre) Funktionen ad hoc zu Design-Absichten und Design-Kompromissen zu erklären. So wird zum Beispiel behauptet, die inverse Netzhaut sei kein Mangel, sondern geradezu erforderlich, um die Energieversorgung des Wirbeltierauges sicherzustellen:
"So ist der Bedarf an Nährstoffen und Sauerstoff für die menschliche Netzhaut deutlich größer als der bei Tintenfischen, was ein komplett anderes Versorgungs- und Entsorgungssystem über die Außenseite und Innenseite der Netzhaut erforderlich macht." (S. 281)
Als ob das bei Greifvögeln, die (entgegen den Cephalopoden) komplett auf eine Vaskularisation der Netzhaut verzichten, nicht erst recht erforderlich und gewährleistet wäre! Warum ein Verlegen der Blutgefäße und versorgenden Zellen hinter die Sehzellebene kein adäquates Sehen ermöglichen sollte, bleibt das Geheimnis des Autors.
Ein anderes Beispiel: Um die verkehrte Brutbeutel-Öffnung der Koalas (Abb. 5) in einen "eleganten Design-Kompromiss" umzumünzen, wird gern das "Kot-Argument" bemüht: Die inverse Öffnung diene dazu, das Junge in die Nähe des Afters der Mutter zu bringen, um es mit dem mütterlichen Kot essenzielle Darmbakterien fressen zu lassen.
Bizarr sind solche Ad-hoc-Hypothesen schon deshalb, weil sie ein Konstruktionsdefizit mit einem anderen weg "erklären": Dass Koalas ihr einziges Nahrungsmittel, die Eukalyptusblätter, nicht selbst verdauen können, ist nämlich alles andere als ein Geniestreich. Das ist "ungefähr so, als wären Löwen allergisch gegen Fleisch" (STEFFENS 2007, S. 17).

Abb. 5 So possierlich der Koala ist, so massiv kumulieren sich bei ihm die Designfehler: Sein Brutbeutel ist nach unten geöffnet. Das Tier kann seine einzige Nahrungsquelle nicht selber verwerten. Zudem muss es den Balanceakt vollführen, gerade so viele Blätter zu futtern, dass es nicht verhungert. Frisst es mehr, erleidet es eine Vergiftung, denn der Koala verträgt die Eukalyptusblätter nur mäßig. In der Konsequenz ist das Tier so geschwächt, dass es täglich 20 Stunden Schlaf benötigt, um nicht an Erschöpfung zu sterben.
Der Schluss auf eine evolutionäre "Bastelei" ist daher naheliegend: Koalas sichern sich durch die Spezialisierung auf (für andere Wirbeltiere unverträgliche) Eukalyptusblätter eine Nahrungsnische ohne Konkurrenz. Das hat den Preis, dass ihr Verdauungssystem von symbiontischen Einzellern abhängt und dass immer nur eine gerade noch verträgliche Menge der Blätter gefressen werden kann. Ein sinnvolles "Design" wäre, den Koalas die Fähigkeit zu geben, selbst passende Enzyme zu produzieren, die den Eukalyptus wirksam entgiften und verdauen. Würde der Brutbeutel nach oben ausgerichtet, wären beide Konstruktionsdefizite beseitigt. Davon abgesehen gäbe es weit sicherere Möglichkeiten, das Darm-Mikrobiom der Mutter auf das Junge zu übertragen.
Großer Beliebtheit erfreut sich auch das "Schnupfen-Argument", das die potenziell lebensgefährliche Kreuzung von Luft- und Speiseröhre in einen genialen Design-Kompromiss verwandeln soll; schließlich könnten wir bei einem Schnupfen durch den Mund atmen und Schleim abhusten (RAMMERSTORFER 2006).
Das Problem ist auch hier, dass es einen Designfehler mit anderen "wegerklärt": Überschießende Schleimbildung (die bei Infekten lebensbedrohlich sein kann) in Kombination mit verkehrt angelegten Abflusswegen der Nasennebenhöhlen (Ostien) birgt weitere Risiken, etwa die Gefahr chronischer Nebenhöhlen-Erkrankungen. Wären die Ostien zweckmäßigerweise nach unten offen, Luft- und Speiseröhre getrennt und durch einen Schließmuskel verbunden, der nur bei Bedarf (Husten, schnelles Atmen) öffnet, wären mehrere Probleme elegant gelöst.
D. Die Strukturen seien aus embryologischer Sicht systemrelevant
Eine Variante des letzten Einwands fordert eine Erweiterung des Blicks auf die Erfordernisse der Embryonalentwicklung (Ontogenese), das heißt auf die Entwicklungsgesetze. Bei biologischen Merkmalen sei zu fragen, ob es ontogenetische Aspekte gebe, die für das betreffende Organ essenziell seien. Sinngemäß lautet das Argument, dass sich Merkmale, die aus technischer Sicht seltsam anmuten, aus entwicklungsbiologischer Sicht als systemrelevant und unverzichtbar entpuppen würden.
Tatsächlich ist die Ontogenese komplizierter ist als das bloße "Verschrauben" von Einzelteilen. Bei technischen Dingen entsteht die Funktion erst am Ende des Fertigungsprozesses. Dagegen muss der Embryo in jedem Stadium seiner Entwicklung funktionieren. Zudem muss er die Voraussetzungen für die jeweils nächsten Entwicklungsschritte schaffen. So werden viele Strukturen etwa als Platzhalter, Induktoren usw. für die korrekte Entwicklung darauffolgender Merkmale und Entwicklungsstadien benötigt. Daher sei es vorstellbar, dass ein Designer kurios anmutende Strukturen in Kauf nahm, weil ihr Fehlen gravierende Probleme an anderer Stelle aufwerfen würde:
"Der typische Aufbau der Netzhaut des Menschen resultiert aus einer in der frühen Embryonalentwicklung beginnenden Interaktion von neuronalen und oberflächlichen embryonalen Gewebeanteilen. Beide Anlagen liefern nicht nur das Zellmaterial für die Ausbildung der Netzhaut. Sie sind eingebunden in ein komplexes Wechselspiel von Induktionen und Hemmungen ontogenetischer Wachstums- und Differenzierungsprozesse im Reigen zahlreicher anderer regulativer Mechanismen. Diese führen u. a. zur Ausbildung der Hilfsstrukturen des Auges wie der Linse, dem Glaskörper oder der Regenbogenhaut …
Auch blinde Fische ohne funktionsfähige Augen im ausgewachsenen Stadium benötigen für die normale Entwicklung ihres Gehirns und des Gesichtsschädels in der Ontogenese Augenanlagen. Argumente des 'unintelligenten Designs' erlangen häufig durch die bewusste Ausblendung der Ontogenese von Organen und Strukturen ihre Plausibilität. Daraus resultiert zwangsläufig eine fehlende Wertschätzung der systemrelevanten formbildenden und funktionellen Aspekte der Organ- und Strukturanlagen, welche weit über ihr zuweilen eigenartig anmutendes Erscheinungsbild (z. B. langer Verlauf der rückläufigen Kehlkopfnerven bei der Giraffe) im ausgebildeten Zustand hinausgehen." (S. 282)
Offenbar übersieht der Autor, dass seine Ausführungen gerade die evolutionäre Argumentation stärken: Die Einbettung in ein komplexes System von Wachstums- und Differenzierungsprozessen ist ja exakt der evolutionsbiologische Grund, warum viele Merkmale "mitgeschleppt" werden, auch wenn sie physiologisch nutzlos wurden! Der Autor erklärt dies am Beispiel blinder Höhlenfische, die ihre Augen behalten. Die naheliegenden (proximaten) Ursachen, ihre Funktion als Induktoren, Platzhalter usw., verhindern eine nachträgliche evolutive Entfernung aus dem Bauplan. Aber die proximaten Ursachen beantworten nicht die Frage nach den ultimaten Faktoren (historischen Determinanten der Entwicklungsprozesse), die uns die Frage beantworten: "Warum sind die Entwicklungsgesetze gerade so, wie sie sind, und nicht anders?"
Aus evolutionärer Sicht ist die Erklärung offensichtlich: Nicht-zielgerichtete Prozesse können das Geflecht historisch gewachsener Zwänge nicht entwirren. Strukturen, einmal in die Ontogenese eingebettet, können nicht mehr grundlegend umgekrempelt, nur noch modifiziert werden. Aber wie will man das mit Geist und Intelligenz erklären, mit Prozessen also, die in der Lage sein sollen, die Merkmale planvoll, zielgerichtet und frei zu kombinieren? Wer Arten oder "Grundtypen" hervorbringt, braucht keine Rücksicht darauf zu nehmen, wie die Entwicklungswege bei anderen Grundtypen aussehen und kann in jedem Fall die optimalen, direkten Entwicklungswege konstruieren.
Wenn es dem superintelligenten Konstrukteur schon gefiel, Giraffen-Embryonen transitorische Elemente hinzuzufügen, die zu einem "Fisch"-Bauplan passen, weshalb hat er das Herz nicht in situ entstehen lassen, statt es in den Brustkorb absteigen zu lassen, sodass es den rückwärtigen Kehlkopfnerv auf ganzer Länge mitnimmt?4) Warum ließ er nicht die rechte Arteria subclavia aus der Aorta distal des Abgangs der A. subclavia sinistra entspringen, sodass der rechte Rekurrens nicht mehr die A. subclavia umschlingt? Diese anatomisch seltene Variante des Nervus laryngeus non recurrens existiert tatsächlich und ist harmlos (HERMANN 2010, S. 131).
Die vermutlich eleganteste Lösung würde darin bestanden haben, die Zielweisung des Nervs durch Modifikationen chemotaktischer Signale so zu verändern, dass der Nerv direkt auf sein Zielorgan zu wächst. Denn Wachstumskegel von Nervenfasern orientieren sich an "diffusiblen" Signalen, die von ihren entfernten Zielen ausgesendet werden. Möchte der Autor behaupten, solche Lösungen würden sich verbieten?
Wenn der Planer ferner darauf Wert legte, in der Embryonalentwicklung hochentwickelter Wirbeltiere auf die archaischen Signalwege von Schädellosen zurückzugreifen, weshalb hat er die Sehzellen nicht in den Augenbecher einwandern lassen, statt sie ins Innere des Neuralrohrs zu verlegen, von wo aus sie später in verkehrter Position aus dem Hirn quellen? Alternativen gäbe es genug. Der Augenbecher des Oktopusses etwa quillt nicht aus seinem Gehirn, sondern entsteht durch Einstülpung der embryonalen Außenhaut. Dann versorgt ihn das Gehirn mit Nervenzellen.
Wir sehen, dass die Probleme nicht konstruktionsbedingt sind. Das Problem ist, dass Embryos "unnötig erscheinende Reminiszenzen an die evolutive Vergangenheit" zeigen (MÜLLER & HASSEL 2018, S. 506). Auf die Frage, warum sich der hypergeniale Planer, statt konstruktive Lösungen anzugehen und neue Signalsysteme zu installieren, historisch gewachsenen Entwicklungszwängen beugt, gibt ID keine auch nur halbwegs überzeugenden Antworten. Die einzig sinnvolle Erklärung lautet: Evolution.
E. Intelligent Design im Rahmen gemeinsamer Abstammung
Um das Problem der Designfehler zu umschiffen, wird gelegentlich folgender Einwand bemüht:
"Der Designer könnte sich dafür entschieden haben, im Rahmen eines historisch limitierten Evolutionsprozesses zu arbeiten. Dies hätte dieselben Einschränkungen im Design zur Folge, wie aus Sicht der etablierten Evolutionsbiologie. Die Kombination aus Intelligent Design und Evolution kann dazu führen, dass ID-Befürworter erwarten, dass das Leben in der von den Evolutionisten beschriebenen Weise suboptimal ist. Aus dieser Perspektive ist schlechtes Design ein Argument für ein evolutionäres Design-Verständnis, weniger ein pauschaler Einwand gegen Design-Argumente." (KOJONEN 2016, S. 157; ins Deutsche MN)
Diese paraevolutionäre Design-Vorstellung (Abb. 6) ist jedoch ein Sonderfall, da sie sowohl die transspezifische Evolution ("Makroevolution") als auch die gemeinsame Stammesgeschichte der Arten zugesteht. Daher kann sich, wie KOJONEN ergänzt, nur eine Minderheit der ID-Vertreter dieses Einwands bedienen. Den WORT-UND-WISSEN-Autoren steht er nicht zur Verfügung, denn damit würden sie weit von ihrer Vorstellung einer technisch intelligenten, biblischen Schöpfung und deren Grundtypen-Modell abrücken. Außerdem zielen die Bemühungen von Paraevolutionisten nicht auf die Destruktion der Evolutionstheorie, sondern auf deren vitalistische "Bereicherung" ab (MAHNER 1986, S. 84).
Im Übrigen führt das Argument in ein auswegloses Dilemma: Wer so weit geht zu behaupten, dass die Einschränkungen in einem intelligenten Designprozess und die historischen Limitierungen nicht-zielgerichteter Evolution dieselben seien, sodass sie zu genau den gleichen Konstruktionsdefiziten führten, zieht sich auf eine Position zurück, die keinen Erklärungs(mehr)wert gegenüber der Evolutionstheorie genießt. Ein solcher Design-Ansatz ist faktisch inhaltsleer: Wenn wir bis hinab in die "Fisch"-Verwandtschaft ontogenetisch limitiert sind, gibt es für Intelligent Design keinen Spielraum, der über das hinausgeht, was natürliche Evolutionsmechanismen auch zu leisten imstande sind.
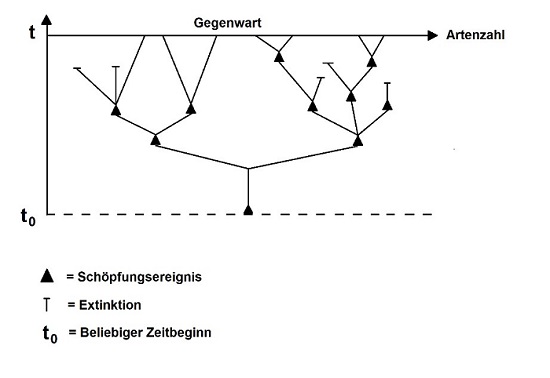
Abb. 6 Paraevolutionäre Vorstellung, wonach im Rahmen der stammesgeschichtlichen Entwicklung der Arten die Verzweigungen von Stammbäumen auf die direkte Intervention eines Schöpfers zurückgehen. Separate "Grundtypen" wurden nicht erschaffen. Nach MAHNER (1986, S. 9).
Anders gesagt: Die paraevolutionäre Design-Idee ist keine prüfbare naturwissenschaftliche Hypothese, sondern eine metaphysische Deutung des Evolutionsgeschehens. Wie soll man untersuchen, ob z. B. die Millionen Jahre zurückliegende Aufspaltung einer Klade auf eine übernatürliche Intervention zurückging? Wo immer wir Artaufspaltungen (Artbildung) direkt beobachten können, hat sie natürliche Ursachen. Es ist vernünftig anzunehmen, dass das auch in den Fällen so war, die wir nicht direkt untersuchen können.
Wer also die Behauptung verteidigen möchte, dass "Intelligenz und Geist" mehr zu leisten imstande seien als "ungelenkte" Prozesse, kommt nicht am Problem der Designfehler vorbei. Es wäre ja willkürlich anzunehmen, dass ein Designer die Möglichkeiten der Evolution immer dann weit in den Schatten stellte, wo er scheinbar Elegantes erschuf, und immer dann dem Potenzial natürlicher Mechanismen verhaftet blieb, wo die banalen Fehler, die unreparierbar verbauten Kompromisslösungen und die Kontingenz gemeinsamer Abstammung sichtbar werden.
Wir sehen also, dass die paraevolutionäre Vorstellung nur eine weitere (nicht prüfbare) Ad-hoc-Annahme im Kabinett der Beliebigkeiten der Intelligent-Design-Verfechter verkörpert. Sie unterstreicht den Vorwurf, dass sich mit einem wie auch immer gearteten Design-Ansatz jeder x-beliebige Sachverhalt "erklären" lässt, sofern nur passende Zusatzannahmen über die Vorgehensweise des Designers getroffen werden.
F. Der Ratschluss des Schöpfers ist unergründlich
Am Ende der (erfolglosen) Ad-hoc-Immunisierung steht immer der Rückzug auf die fehlende menschliche Urteilskraft und auf den unergründbaren Ratschluss des göttlichen Konstrukteurs:
"Dawkins’ anthropomorpher und unintelligenter Designer ist nichts weiter als ein jämmerlicher Strohmann, geboren aus menschlicher Überheblichkeit und Unwissenheit … Dawkins nutzt die menschliche Rationalität, um an Gegenständen der Natur das schlechte Handwerk eines Designers oder Schöpfers nachzuweisen ('unintelligentes Design'). Das gelingt aber nur, wenn er dem kritisierten Konstrukteur – wie oben gezeigt- eine niedrigere Intelligenz als dem Menschen unterstellt. Der Option, möglicherweise einem Gott gegenüber zu stehen, der über ein unendlich höheres Maß an Intelligenz verfügt als der Mensch (wie vor allem im biblischen Zeugnis belegt), ist sich Dawkins offenbar nicht im Geringsten bewusst." (S. 286)
Offenbar fühlt sich der Autor durch die sinngemäße Behauptung des Biologen Richard DAWKINS provoziert, dem mutmaßlichen Designer mangele es eklatant an Verstand. Allerdings zielt DAWKINS' Argument gar nicht darauf, einem übernatürlichen Designer Dummheit zu unterstellen. Er will die Behauptung, es gäbe diesen Konstrukteur, ad absurdum führen.
Der Rückzug auf das "biblische Zeugnis" zeigt, wie der Autor im Kreis argumentiert. Nicht nur, dass die These, die Intelligenz des Designers sei "unendlich höher" als die des Menschen, das zu Beweisende schon voraussetzt. Sie ist auch ein argumentatives Passepartout, ein Immunisierungs-Joker, mit dem man auch die offensichtlichste Pfuschlösung dem unergründlichen Ratschluss des Schöpfers anheimstellen kann. Doch das "Argument" hat schwerwiegende Nebenwirkungen: Wer dem Menschen in dieser Frage kein Urteil zutraut, spricht ihm auch die Fähigkeit ab zu beurteilen, ob biologische Systeme besser mit Evolution oder "Planung" harmonieren.
Anders gesagt: Wenn das mutmaßliche göttliche Schöpfungshandeln unser menschliches Verständnis absolut übersteigt, warum kann es dann nicht ebenso gut (eigentlich viel besser) in einem gewaltigen Prozess der Evolution bestehen, der all die Eigenschaften hat, die ihm Biologen zuschreiben? Die Advokaten des "intelligenten Designers" müssen sich schon entscheiden, ob Menschen die Qualität (bzw. Intelligenz) des angeblichen Designs beurteilen können oder nicht. Wenn ja, dann ist der Rückzug auf unergründliche, übernatürliche Ratschlüsse sinnlos. Wenn nein, ist ihre gesamte Argumentation sinnlos und sie können sich ebenso gut an die biologischen Lehrbücher halten.
Einen raffinierteren Versuch zu argumentieren, uns fehle für eine angemessene Beurteilung von Designfehlern derzeit das erforderliche entwicklungsbiologische Wissen, unternimmt RAMMERSTORFER (2004). Doch auch hier gilt, dass mit der Frage, ob eine entwicklungsbiologische "Urteilsbasis" existiert, das Planmäßigkeits-Argument steht und fällt: Existiert diese Basis nicht, können wir auch nicht beurteilen, wie speziell und zielorientiert die Teile eines Systems zusammengefügt sind und wie gut sie einer Zweck-Mittel-Beziehung entsprechen (ähnlich argumentiert KOJONEN 2016, S. 159).
Auf der Basis derzeitigen Wissens sind allerdings durchaus Urteile darüber möglich, ob Designfehler vorliegen und ob es bessere ontogenetische Alternativen gibt. Anhand des rückwärtigen Kehlkopfnervs und des Wirbeltierauges haben wir Beispiele erörtert. Künftige Forschungsergebnisse könnten dieses Urteil revidieren; einstweilen aber gilt, dass nicht-zielgerichtete Evolution die derzeit beste und völlig befriedigende Erklärung darstellt.
Literatur
BRINGMANN, A. & REICHENBACH, A. (2001) Role of Muller cells in retinal degenerations. Frontiers in Bioscience 6, E72-92.
BRINGMANN, A. et al. (2006) Muller cells in the healthy and diseased retina. Progress in Retinal and Eye Research 25, S. 397-424.
EBERHARDT, C. (2011) Die Bedeutung der Müllerzelle für die Pathogenese der equinen rezidivierenden Uveitis und Etablierung einer equinen Müllerzelllinie. Inaugural-Dissertation zur Erlangung der tiermedizinischen Doktorwürde der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität München.
FRANZE, K. et al. (2007) Müller cells are living optical fibers in the vertebrate retina. PNAS 104, 8287-8292.
FRANZE, K. & GROSCHE, J. (2008) Müller-Zellen in einem anderen Licht. Biospektrum 14, S. 701-703.
HASS, H. (1979) Wie der Fisch zum Menschen wurde. Die faszinierende Entwicklungsgeschichte unseres Körpers. C. Bertelsmann, München.
HERMANN, M. (2010) Schilddrüsenchirurgie. Springer-Verlag, Wien, New York.
JUNKER, R. & WIDENMEYER, M. (2021) Schöpfung ohne Schöpfer? Eine Verteidigung des Design-Arguments in der Biologie. SCM Hänssler, Holzgerlingen.
KOJONEN, E. R. V. (2016) The Intelligent Design debate and the temptation of scientism. Routledge.
LU, Y. B. et al. (2006) Viscoelastic properties of individual glial cells and neurons in the CNS. PNAS 103, S. 17759-64.
MAHNER, M. (1986) Kreationismus. Inhalt und Struktur antievolutionistischer Argumentation. Pädagogisches Zentrum, Berlin.
MAHNER, M. & BUNGE, M. (2000) Philosophische Grundlagen der Biologie. Springer, Berlin.
MÜLLER, W. A. & HASSEL, M. (2018) Entwicklungs- und Reproduktionsbiologie des Menschen und bedeutender Modellorganismen. 6. Auflage. Springer, Berlin.
MYERS, P. Z. (2010) More creationst misconceptions about the eye.
RAMMERSTORFER, M. (2003) Eine Theorie zur Signalerkennung und ihre (möglichen) Folgen. Webarchiv, Memento vom 23.07.2021
RAMMERSTORFER, M. (2004) Nervus laryngeus recurrens - suboptimal? Webarchiv, Memento vom 23.07.2021.
RAMMERSTORFER, M. (2006) Nur eine Illusion? Biologie und Design. Tectum-Verlag, Marburg.
RIEDL, R. (1984) Die Strategie der Genesis. Piper-Verlag, München.
STEFFENS, D. (2007) Tierisch! Expeditionen an den Rand der Schöpfung. Reinbek, Hamburg.
STEINBRÜCK, I.; BAUMHOER, D. & HENLE, Ph. (2008) Intensivkurs Anatomie. Urban & Fischer Verlag, München.
WILLIAMS, R. (2006) Unintelligent Design: Why god isn't as smart as she thinks she is. Crows Nest, N.S.W.
Fußnoten
[1] Dieses Beispiel verdeutlicht, warum Intelligent Design (ID) nicht in der Lage ist, ein alternatives Erklärungsparadigma anzubieten. Zwar wird die Rekurrens-Schlinge samt Reminiszenz an den "Fisch"-Bauplan ad hoc als Design-Kompromiss zu deuten versucht (siehe unten). Verliefe aber die Entwicklung der Säugetiere von der molekularen bis zur anatomischen Ebene vollkommen anders als die der Fische, so dass auch der Kehlkopfnerv zielgerichtet zum Kehlkopf führen würde, ließe sich dies erst recht als Hinweis auf das kreative Genie eines superintelligenten Schöpfers deuten, der die Merkmale "frei" kombiniert habe, ohne an die Kontingenz gemeinsamer Abstammung gebunden zu sein! Die akademische Evolutionstheorie hingegen könnte einen solchen Befund nicht erklären, ja sie wäre wohl gar nicht erst entstanden. Wir sehen also, dass Beliebiges unter das "Design"-Paradigma subsumiert werden kann. Aber ein Paradigma, das problemlos alles erklärt, erklärt leider gar nichts.
[2] Zwar könnte man "Design" so unspezifisch halten, dass jedweder Pfusch damit kompatibel wäre. Aber dann bräuchte man sich über diejenigen Design-Indizien, die auf die "Zielorientierung" bzw. Sinn-, und Zweckhaftigkeit biotischer Merkmale abheben, gar nicht mehr zu unterhalten.
[3] Hier könnten die Autoren versucht sein, eine Analogie mit der Technik zu bemühen und zu entgegnen, dass auch ein stotternder, strukturell defizitärer Motor erkennbar ein Beispiel für Planmäßigkeit ("Design") wäre. Dieser Vergleich ginge jedoch am Sachverhalt vorbei, da hier unausgesprochen so viel Hintergrundwissen einflösse, dass wir selbst einen Haufen Metall-Schrott noch als "gemacht" erkennen würden. In der Biologie haben wir dieses Wissen nicht, im Gegenteil: Wir wissen, dass Organismen evolvieren und dass defizitäre Strukturen (maximal optimiert zwar, aber trotzdem zweitklassig) exakt den Erwartungen von Evolutionsprozessen entsprechen, die keinen Plan und kein Ziel kennen.
[4] Da sich die Herzanlage aus zugewanderten Angioplasten und Myocardvorläuferzellen bildet, die wie Amöben über den Vorderdarm kriechen und sich zu den Herzfeldern zusammenschließen, spricht nichts gegen diese Möglichkeit. Wie weit sie nach vorn wandern, wird durch angiogene Faktoren bestimmt.
Autor: Martin Neukamm & Hansjörg Hemminger
